|
| Foto: Magnus Lundgren |
Negrito
Etmopterus spinax (Linnaeus, 1758)
(es. Negrito, cochino; gal. Gata, gata negra, crabudo, demo; ing. Velvet belly; port. Lixinha da fundura)
Orden: Squaliformes
Familia: Etmopteridae
Para celebrar estas primeras Navidades de
Tiburones en Galicia, nada mejor que traer un tiburón que brilla con luz propia no sólo en el sentido figurado. Es decir: un tiburón que da luz, un tiburón luminoso de verdad, que tanto nos puede servir para coronar el árbol de Navidad como para guiar a SSMM los Reyes Magos de plástico hasta la entrada de la gruta que casi todos los agnósticos y ateos tenemos montada en el salón. En definitiva, una bombilla con aletas y dientes... pero infinitamente más auténtica, ecológica y bella que las que adornan las calles del país.
El negrito (
Etmopterus spinax) es una de las cerca de 50 especies que conforman el género
Etmopteridae, caracterizadas, entre otras cosas, por la presencia más o menos masiva de fotóforos
—órganos productores de luz— en diversas zonas de su cuerpo, particularmente en la superficie ventral. En nuestras aguas contamos con tres de estos tiburones, los tres pertenecientes al mismo género,
Etmopterus. Además del
spinax tenemos el
E. pusillus o tollo lucero y
E. princeps o tollo raspa. Las tres especies bastante parecidas, difíciles de identificar. Las diferencias morfológicas pueden bastante sutiles y algunas imposibles de detectar a simple vista, como por ejemplo los dentículos dérmicos.
 |
La superficie ventral, más oscura, revela la presencia de fotóforos (foto: CEMMA).
|
 |
| Superficie ventral "encendida". |
El tema de la bioluminiscencia lo vamos a tratar en otros artículos. Por el momento, baste decir que, lejos de ser una rareza, en realidad es un fenómeno que se da en alrededor del 10% de las especies conocidas, además de ser un fenómeno digamos que normal entre las criaturas que habitan las aguas profundas.
La generación de luz en los etmoptéridos está controlada mediante la actividad hormonal, en concreto de la prolactina y la melatonina [véase Bioluminiscencia I: Los fotóforos]. El objetivo, según la teoría más extendida, es el camuflaje —vista desde abajo, la superficie ventral iluminada se difuminaría contra la claridad de la superficie, como el contrasombreado de los tiburones de aguas más someras— y también, tal vez, la comunicación entre individuos en la oscuridad de las aguas profundas [Bioluminiscencia II: Funciones].
 |
Foto: CEMMA.
|
Descripción: Cuerpo cilíndrico, grueso y alargado, con dos largas espinas delante de las dorsales. La primera es claramente más corta que la segunda. Las hendiduras branquiales son muy pequeñas, prácticamente de la misma longitud que los espiráculos (1/3 de la longitud del ojo). Morro corto y romo, de longitud similar a la anchura de la cabeza. La boca es recta en los ejemplares adultos, mientras que en los jóvenes está un poco arqueada.
El origen de la primera dorsal se encuentra detrás del borde posterior de las aletas pectorales. La segunda dorsal es claramente mayor que la primera (casi el doble), y se origina sobre la axila de las pélvicas. Las aletas pectorales son pequeñas y trapezoidales. El lóbulo inferior de la caudal está poco definido. Como todos los Squaliformes, el negrito carece de aleta anal.
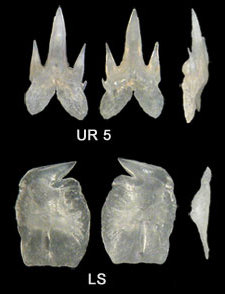
Los dentículos dérmicos son de base muy estrecha y presentan una cúspide central muy larga, curvada y flexible.
 |
Arriba izq., dentículos con 28x (foto: Albert Lleal); abajo dcha., dentículos con 221x (foto: Albert Lleal), sobre una excelente fotografía de Andy Murch, sharksandrays.com.
|
En cuanto a su librea, el color varía desde el negro azulado al gris pardusco, con la zona ventral muy oscura o negra. Presentan también unas bandas oscuras sobre las pélvicas y la caudal.
Dentición: Dimorfismo dentario: los dientes superiores tienen entre 3 y 5 cúspides y se encuentran espaciados y dispuestos en 2 o 3 hileras funcionales; los inferiores, en cambio, presentan una sola cúspide, muy abatida, y están imbricados y dispuestos en una única hilera funcional formando como una cuchilla. Sin sinfisarios.
Talla: Los datos sobre tallas varían un poco según el trabajo que consultemos. No sabemos a ciencia cierta el porqué, si es debido a factores geográficos o ambientales, a la utilización de diferentes sistemas de muestreo, o bien
a la propia actividad pesquera: se sabe que el tamaño de las especies disminuye en las zonas donde la presión pesquera ha sido mayor. Al nacer miden de 12 a 14 cm, aunque en el Adriático se ha informado de neonatos de 8-10 cm. La longitud total máxima registrada fue de 60 cm
—alcanzados, como ya os imagináis, por una hembra
—, aunque por lo general estos bichos raramente superan los 45 cm.
Lo que sí podemos afirmar con certeza es que el
Etmopterus spinax es el tiburón más pequeño de Galicia.
Reproducción: Vivíparo aplacentario (ovovivíparo), con camadas de entre 6 y 20 crías.
 |
| Foto: Fumio Nakagawa (1) |
Existen divergencias en cuanto a la talla de madurez del negrito. Compagno la sitúa en un genérico y
unisex 33-36 cm; otros, por su parte, extienden el abanico y apuntan
a >30-46 cm para las hembras y 25-38 cm para los machos
—los 25 cm corresponden a un ejemplar capturado en el Algarve
—. Lo que cada vez se hace más evidente en todos estos trabajos es que las
hembras maduran más tarde que los machos, y con tallas superiores. En parte esto se debe a la necesidad de alcanzar un tamaño suficiente para albergar a sus retoños y, a la vez, poder dedicar a tareas reproductivas la energía empleada para el propio crecimiento. Por ese motivo las hembras tienen también un hígado más grande. (2)
Como ocurre con la gran mayoría
—por no decir la totalidad— de los tiburones de aguas profundas, la tasa reproductiva del negrito es sumamente baja: baja fecundidad y ciclo reproductivo bastante largo, estimado entre 2-3 años. A ello hay que añadir que es una especie de crecimiento lento, lo cual implica que tarda bastante más tiempo que los tiburones de aguas más someras en alcanzar la madurez sexual, es decir, en estar preparados físicamente para la reproducción.
Un dato ilustrativo: en el Atlántico oriental la edad máxima de los machos y la hembras muestreados fue de 8 y 11 años, respectivamente; pues bien, lo que se observó fue que la madurez sexual llegaba habiendo alcanzado, respectivamente, en torno al 49,6% y 42,5% de esa edad máxima (3).
O sea. Teniendo todos estos parámetros en cuenta, con un poco de suerte una hembra de negrito podrá reproducirse un máximo de 3 veces en toda vida, produciendo, si es una atleta campeona, un máximo de 60 crías. Ahora, que cada cual saque sus conclusiones.
Dieta: A base de pequeños peces demersales, calamares, crustáceos y otros organismos del fondo.
Hábitat y distribución: El negrito es un tiburón demersal bentónico del talud superior y la plataforma continental exterior. Su rango batimétrico es amplio, desde los 70 hasta los 2000 m, si bien principalmente entre los 200 y los 500 o 600 m, según la zona geográfica. Por ejemplo, en las Azores no se han registrado capturas por debajo de los 1500 m, mientras que en el Mediterráneo se les ha llegado a capturar a 2450 m. Posiblemente estas diferencias obedezcan a factores hidrográficos como la temperatura o la salinidad, o a factores relacionados con la actividad trófica, etc.(4)
Igualmente, al menos en algunas zonas geográficas parece detectarse una segregación batimétrica por tamaño y sexo: los ejemplares más grandes, las hembras, se encuentran a mayor profundidad.
 |
| (Fuente: Wikipedia) |
Se encuentra en aguas frías a tropicales del Mediterráneo y el Atlántico
oriental desde Islandia hasta Costa de Marfil y Nigeria, pasando por
las Azores, Canarias, Cabo Verde, y puntos de Sudáfrica.
En Galicia es una especie común. Cito textualmente de la guía de Rodríguez Villanueva y X. Vázquez (5):
En Galicia ten sido citado en fondos maiores de 200 m., dende Fisterra a Estaca de Bares e dende o alfoz do Miño ata Estaca de Bares.
 |
| (Foto: thejump.net) |
Pesca y estatus: El negrito no es una especie comercial. Su carne se consume, pero en nuestra zona forma parte de las capturas accidentales de artes de fondo como el palangre o el arrastre, y se destina a harina de pescado, si bien por lo general se descarta. Así ocurre en la flota que faena al sur de Portugal, donde las capturas son abundantes por parte de los arrastreros que andan a la gamba (
Parapenaeus longirostris), la cigala (
Nephrops norvegicus), o el listado o gamba roja (
Aristeus antennatus); o los palangreros que se dedican a la pescadilla (
Merluccius merluccius), el congrio (
Conger conger) y el mero (
Polyprion americanus), etc. Una parte de los negritos que se descartan siguen todavía vivos, pero tan dañados que sus posibilidades de supervivencia se ven sustancialmente reducidas. Los índices de CPUE (Capturas por Unidad de Esfuerzo) son más que elocuentes: se calcula que los arrastreros que operan entre los 200 y los 700 m
—muy particularmente en torno a los 500 m
— capturan una media de 15,74 especímenes por hora. (6)
Aunque se trata de una especie relativamente común, su baja tasa reproductiva la hace extraordinariamente vulnerable a la sobrepesca. A esto hay que sumar el tema de segregación espacial por tamaño: todo parece indicar que, al menos en el Atlántico nororiental, las hembras maduras se suelen concentrar a una cota de 600 m, lo cual implica que un solo lance puede cercenar las perspectivas de supervivencia de toda una población.
La Lista Roja de la IUCN con el estatus de
Preocupación menor atendiendo al hecho de que en general las cifras de registros de capturas parecen mantenerse a un nivel constante, es decir, no hay evidencias de un descenso poblacional. No obstante, se establece una situación de excepcionalidad para el Atlántico nororiental, tan intensamente explotado: los estudios científicos han constatado una disminución del 20% en el tamaño de las poblaciones desde los años 70 hasta el 2004, lo cual, unido a la creciente presión pesquera en aguas profundas, le garantiza el estatus de
Casi amenazado.
 |
| (Fotografiado en Noruega por Rudolf Svensen) |
Si
no lo estuviésemos destrozando a velocidad supersónica, nuestro fondo
marino podía parecerse a la ciudad de Papá Noel, repleta de
bichitos raros correteando por todos los rincones y montones de lucecitas chispeando aquí y allá. De aquí a pocos años no quedará
nada.
Sea de ello lo que fuere, que tengáis todos una feliz Navidad junto con todos los vuestros y un feliz año nuevo.
_______________________
(1) En realidad, estos dientes no pertenecen a un Etmopterus spinax, sino, según el dueño de la página donde figura esta fotografía, a un E. pusillus. Los dientes de ambas especies son casi idénticos, de ahí que, a falta de algo mejor, me haya atrevido a daros el cambiazo.
(2) y (4) Ana Aranha et al. "Biological aspects of the velvet belly lantern shark, Etmopterus spinax (Linnaeus, 1758) off the Azores, North East Atlantic." Marine Biology Research, 2009; 5:3, 257-267.
(3) R. Coelho y K. Erzini. "Life history of a wide-ranging deepwater lantern shark in the north-east Atlantic, Etmopterus spinax (Chondrichthyes: Etmopteridae), with implications for conservation." Journal of Fish Biology (2008). 73, 1410-143.
(5) X. L. Rodríguez Villanueva y Xavier Vázquez. Peixes do mar de Galicia (I): Lampreas, raias e tiburóns. Vigo: Xeráis, 1992, p. 81.
(6) Para estos (y otros) datos, ver el trabajo citado de Coelho y Erzini, y la ficha de la especie en la página de la IUCN Red List.